近日,南方科技大学生命科学学院副教授吴柘与助理教授陈曦团队合作在国际知名学术期刊Nature Genetics(IF~39)上发表题为“Distinct chromatin signature in Arabidopsis male gametophyte”的研究论文。该研究建立了一套适用于植物雄配子体细胞的超高灵敏度染色质免疫共沉淀测序(ChIP-seq)方法,并结合染色质开放性测序 (ATAC-seq)以及单细胞转录组测序 (scRNA-seq) 等多组学技术,首次展示了模式开花植物拟南芥雄性生殖细胞系发育及成熟过程中组蛋白修饰H3K27me3及H3K4me3重编程的特征和机制,为理解植物表观遗传信息如何在代际间遗传和重置提供了知识参考。

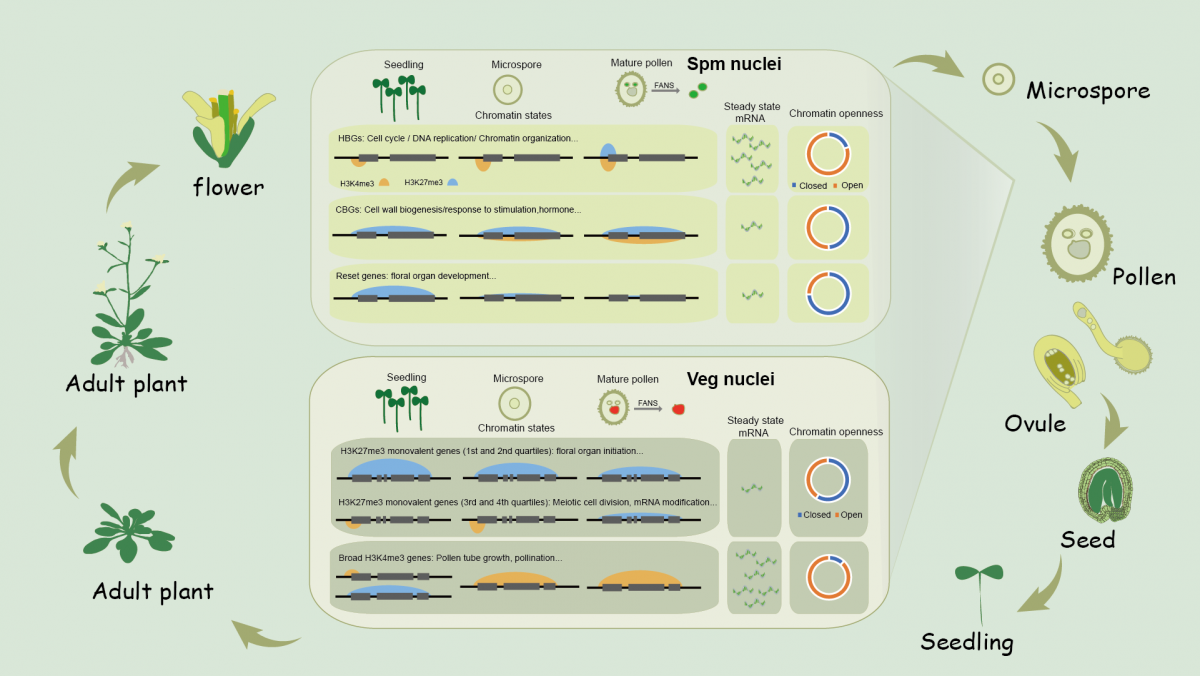
表观遗传信息通常指在染色质上除DNA序列信息之外对中心法则的执行起到促进或者抑制作用的各类修饰,比如DNA甲基化、组蛋白甲基化等。表观遗传信息的存在对细胞命运决定及维持、细胞环境信号响应等方面都至关重要。表观遗传信息虽然经常能经过细胞分裂传递给子代细胞,但科学家对表观遗传信息究竟能在多大程度通过有性生殖传递给后代一直存在争议和讨论。在哺乳动物中,得益于生殖细胞成熟以及受精后的几次重编程过程,一般认为上一代获得的表观遗传信息极少传递给后代。与动物不同,有花植物在胚胎发育的过程中并未分化出生殖细胞,在胚胎发育之后仍将经历一段较长时间的营养生长阶段,在此过程中植物需要不断响应内源及环境信号,随之分化产生生殖器官(花)及生殖细胞。以植物雄性生殖细胞的发生为例:小孢子母细胞由花药孢原细胞分化形成,随后小孢子母细胞经减数分裂产生4个小孢子(microspore,Ms),每个小孢子再经历两次有丝分裂形成一粒包含了两个精细胞(sperm cell, Spm)及一个营养细胞核(vegetative nucleus, Veg)的成熟花粉。不难看出,有花植物精细胞是由体内一小部分体细胞经过几次细胞分裂直接“去分化”形成的。有趣的问题是:亲本的表观遗传信息是如何在体细胞到精细胞的几次细胞分裂事件中被“重编程”的?精细胞及营养细胞中染色质修饰特征是什么?表观遗传信息的重编程如何与精细胞及营养细胞的命运相偶联?此前受实验技术限制,这些重要的科学问题在植物生殖领域一直悬而未决。
该研究综合了此前发表的ChIPmentation技术及native ChIP技术的优点,首次建立了一套适用于植物的超高灵敏组蛋白表观修饰检测技术。借此方法,研究人员首先发现在模式植物-拟南芥的精细胞染色质中存在广泛的染色质二价性(约占80% H3K27me3修饰位点), 即通常认为的抑制型组蛋白修饰(H3K27me3)与激活型组蛋白修饰(H3K4me3)广泛共存于染色质的不同区域。结合此前的报道,染色质二价性广泛存在于哺乳动物的一些多能干细胞中,而在动植物的体细胞中均基本不可见,因此该研究的结果暗示了染色质二价性可能是具有多能性或全能型细胞的一个保守的染色质特征。进一步,根据精细胞稳态下的基因表达水平,具有二价性修饰的基因又能被进一步分为与活跃基因表达相关的HBG(Hot Bivalent Gene, FPKM>= 0.5) 及与沉默基因表达相关的CBG (Cold Bivalent Gene, FPKM<=0.5)两大类。 HBG及CBG分别基于在小孢子时期就已经预先建立的H3K4me3及H3K27me3而建立。在功能上,HBG基因主要富集了DNA复制、修复、细胞分裂等在雄配子体发育过程中所必须的基因,而CBG则主要富集了细胞壁合成和环境刺激响应等在精子发生过程中不被需要但对在受精后可能会需要其表达的基因。通过综合分析,该研究提出了HBG及CBG的可能作用均在于使基因处在一种比较灵活的停顿状态(poised state),为受精后基因表达或抑制程序的启动和运行做准备。

图1:通过Native ChIP-seq结果显示染色质二价性广泛存在于拟南芥精细胞中
植物生殖细胞由体细胞直接“转变”而来,体细胞中建立的表观遗传信息能否及如何传递给生殖细胞一直是个待解之谜?在此背景下,研究人员细致分析了H3K27me3修饰从体细胞、小孢子、到成熟精细胞这一过程中的动态变化。发现大部分体细胞中H3K27me3修饰位点上的修饰水平在精细胞发育过程中存在下调。然而,令人惊讶的是对H3K27me3较为彻底的擦除只存在于包括FLC在内的700多个基因上,这些基因高度富集了对花器官发育及胚胎模式建成等发育过程起重要作用的转录因子。该结果揭示了全表观组水平H3K27me3修饰在植物雄性生殖细胞中重置的逻辑,且进一步暗示,对亲本表观修饰进行局部而非整体的擦除也许就足以保证子代植物正确的发育及环境信号响应。

图2: 组蛋白修饰(H3K27me3)的重置选择性发生在一些对发育起重要作用的基因上
此外,领域内的一种观点认为精细胞中特异存在的组蛋白变体H3.10会通过渗入并替换传统的组蛋白H3.1从而引起H3K27me3在精细胞中的重置。通过在H3.10基因敲除的突变体中研究其精细胞的染色质修饰,该研究发现H3.10对重置体细胞的H3K27me3贡献微弱,数据表明伴随花粉第一次有丝分裂的完成,组蛋白H3.10可能优先渗入并影响一些在生殖细胞(Generative cell, GC)中H3K27me3修饰水平极低的位点,其渗入阻止了在这些位点上进一步建立H3K27me3的可能性;而巧妙的是,上述这些位点正是在营养细胞中需要被建立的位点。因此,H3.10的渗入帮助了精细胞建立起区别于营养细胞的染色质特征,其主要功能不在于帮助重置体细胞的H3K27me3。
高等植物生殖细胞除了在产生方式上与动物存在差别外,它们在形态和受精方式上也存在巨大差异。拟南芥的两个精细胞与一个体积较大的营养细胞核共同存在于同一花粉粒中。在受精过程中,营养细胞通过花粉管萌发,将两个精细胞输送到未受精胚珠附近完成双受精。通过分析营养细胞染色质修饰,该研究发现营养细胞代表了一类高度特化的细胞类型,染色质的修饰状态与营养细胞的功能非常吻合。有数千个位点在小孢子到营养细胞核形成过程中新建了H3K27me3的修饰,其中许多是减数分裂时被需要而在后续过程中被关闭的基因。与此同时,与花粉管萌发,受精等过程高度相关的基因在营养细胞中活跃表达,它们的染色质中则被“宽泛”的H3K4me3修饰覆盖在整个基因区域。此外,与营养细胞核松散的形态相一致,染色质可及性数据(ATAC-seq)显示营养细胞核染色质较精子及体细胞都更为开放。在近着丝粒区域,营养细胞中升高的染色质可及性伴随了H3K27me3在此区域的积累;与此同时,H3K9me2仍然在该区域存在明显富集。此外,不同的组蛋白修饰状态可以对应到不同的染色质可及性,显示染色质松散程度并非简单反应了基因的表达水平,而更可能关联到基因的转录潜能。
本项目由吴柘课题组和陈曦课题组通过紧密合作完成,吴柘课题组研究副教授朱丹灵完成了本论文绝大部分实验,陈曦课题组博士生闻懿完成了绝大部分对高通量测序数据的处理和分析。朱丹灵和闻懿为该论文共同第一作者。吴柘与陈曦为论文共同通讯作者,南科大为论文第一单位。美国罗格斯大学学者郑海燕在蛋白质谱分析中提供了关键数据。吴柘课题组博士生姚婉玥、周思娴、张琦琦也参与了本项研究。北京大学教授瞿礼嘉在论文科学问题凝练及数据解释方面提供了宝贵指导。该研究得到了国家自然科学基金,广东省科技厅创新创业团队项目,广东省普通高校重点实验室以及深圳市科创委基金的资助。
论文链接: https://www.nature.com/articles/s41588-023-01329-7
供稿:生命科学学院
通讯员:付文卿
主图:丘妍
编辑:朱增光





南方科技大学微信

南方科技大学视频号

南方科技大学抖音号

南方科技大学快手号

南方科技大学头条号

南方科技大学南方+